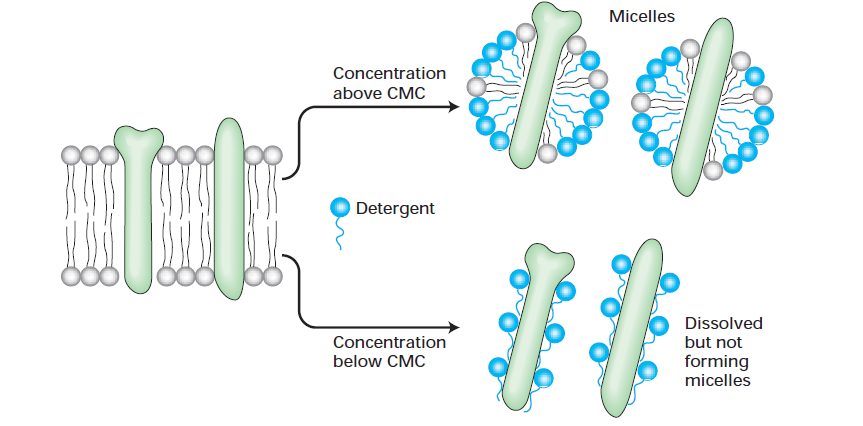
انواع حالات برهمکنش دترجنت ها با پروتئین های اینتگرال غشایی
+ نوشته شده در شنبه ششم اسفند ۱۳۹۰ ساعت 15:8 توسط امیر اکبری آرمند
|


The first two steps in plasmalogen biosynthesis take place in peroxisomes by the peroxisomal enzymes DHAP-AT and ADHAP-S. The third step may occur both in peroxisomes and in the endoplasmatic reticulum (ER), since AADHAP-R has been found in both subcellular compartments. All remaining steps occur in the ER.
هیستونها فراوانترین پروتئینهای متصل به DNAیوکاریوتی هستند.به طور معمول 5هیستون فراوان موجود در سلولهای یوکاریوتی H1/H2A/H2B/H3/H4میباشند.هیستونهای H2A/H2B/H3/H4را کور هیستون مینامند که هسته پروتئینی تشکیل داده و DNAنوکلئوزومی در اطراف آن میپیچد.همانطور که متوجه شدید H1 جزء کور هیستون نمیباشد.این هیستون به DNAلینکر متصل میشود از این رو به H1هیستون لینکر میگویند.هر 4هیستون کور هیستون به مقدار مساوی در سلول یافت میشوند ولی فراوانی H1نصف سایر هیستونهاست.این موضوع با این یافته که به هر نوکئوزوم تنها یک مولول H1(و دو کپی از هر کور هیستون)متصل میشود.
با توجه به ارتباط نزدیک هیستونها با بار منفی مولکول DNA،آنها دارای آمینو اسیدهایی با بار مثبت زیاد میباشند.بیش از 20% باقی مانده ها در هر هیستون ،لیزین و آرژنین میباشد.کور هیستونها پروتئینهای نسبتا کوچکی بوده و وزنی بین 11-15 کیلو دالتون دارند،درحالیکه H1تقریبا 20کیلو دالتون است.
هسته پروتئینی نوکلوزوم،یک ساختار دیسکی شکل(دیسکوئید)داشته و تنها در حضور DNAبا نظم خاصی تجمع میابد.در غیاب DNA،کور هیستونها در محلول تشکیل دسته های حد واسطی می نمایند.یک ناحیه حفظت شده در هر کور هیستون به نام دومین شکل گیری هیستون(HISTON FOLD DOMAIN)تجمع این حدواسطهای هیستونی را موجب میشود.دومین شکل گیری هیستون از سه ناحیه آلفا هلیکس تشکیل شده که توسط 2 لوپ کوتاه بی شکل از یکدیگر جدا شده اند.در هر حالت ،دومین شکل گیری هیستون،تشکیل هترودایمرهایی از جفتهای اختصاصی هیستونها به صورت اتصال سر به دم را موجب میشوند.ابتدا H3وH4هترودایمر داده،سپس با دو مولکول یکی H3و دیگری H4با هم تشکیل تترامر میدهند.در مقابل H2AوH2Bدر محلول هترو دایمر میدهند(نه تترامر)
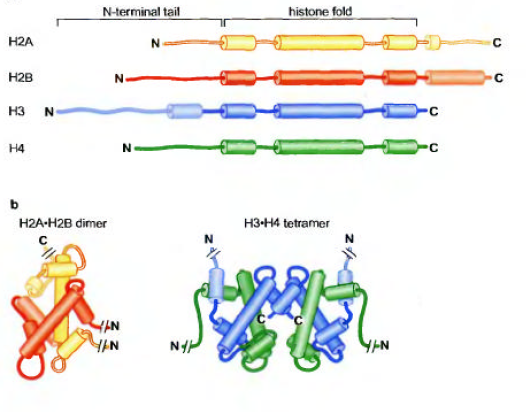
تجمع نوکلئوزوم شامل اتصال منظم این واحدهای ساختاری با DNAمیباشد.ابتدا تترامرH3وH4به DNAمتصل میشود،سپس دو دایمر H2AوH2Bبه کمپلکس H3-H4-DNAمتصل و نوکلئوزوم نهایی را تشکیل میدهند.
نکته1:تترامرهای H3-H4و دو دایمر H2A-H2Bهر کدام با ناحیه مشخصی از DNAدر نوکلئوزوم میانکنش میدهند.به طوریکه اگر نوکلئوزوم را به صورت صفحه ساعت توضیح دهیم،تترامرH3-H4نیمه بالایی اکتامرهیستونی را تشکیل میدهد،در مقابلH2A-H2Bقسمت پایین اکتامر هیستونی را تشکیل میدهد.
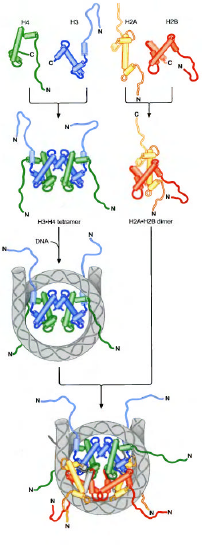
نکته2:هیستونH1،با LINKER DNAاینترکشن داده و اتصال DNAبا نوکلئوزوم را محکمتر میکند.
منبع:کتاب ژنتیک مولکولی واتسون
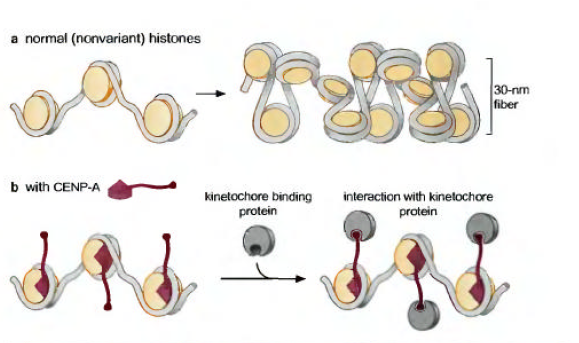
CENP-Aهیستون متفات دیگری است که در نوکلئوزومهای موجود در DNAسانترومری دیده میشود.در این ناحیه کروموزومی،CENP-Aدر نوکلئوزوم،جایگزین H3 است.این نوکلئوزومها در کینه توکور(با استناد به کتاب دسل ،این نوکلئوزوم به صفحه کینه توکور داخلی وارد میشود)شرکت میکنند.CENP-Aدر مقایسه با H3دارای ناحیه دم N-انتهایی بزرگتر و در سایر جنبه ها مشابه هیستونها میباشد.بنابراین شرکت CENP-Aبعید است که هسته ساختار نوکلئوزوم را تغییر دهد.در مقابل احتمالا دم کشیده آن جایگاههای جدیدی برای اتصال سایر اجزای پروتئینی کینه توکور به وجود می آورد.همگام با این نظریه فقدان CENP-A ،مانع اتصال اجزای کینه توکور با سانترومر میشود.
منبع:کتاب ژنتیک مولکولی واتسون
لازم میدانم بگویم CENPها از پروتئینهای سانترومر هستند و شامل CENP-A/CENP-B/CENP-Cهستند.در ضمن کینه توکور هم 4بخش است:1-صفحه کینه توکور داخلی/2-ناحیه ای که بین صفحه کینه توکور داخلی و خارجی است/3-صفحه کینه توکور خارجی/4-فیبروس کورنا
منبع:کتاب دسل


Figure. PCNA
organises many steps at the replication fork. Here it is shown interacting with the replicative polymerases on both the leading and lagging strands, and on the lagging strand also coordinating Okazaki fragment processing by Fen-1 and Ligase1.
عملکرد SRP در پروکاریوت ها با یوکاریوت ها متفاوت است
از تمام عزیزانی که حتی با اندازه ی یک سلام بر گردن من حق دارند/تقاضامندم قبل از شروع ماه مبارک رمضان من را حلال کنند.
السلام علی عبادالله الصالحین
التماس دعا
 با عرض سلام و احترام
با عرض سلام و احترام